PROPÓSITOS:
Se estudiaran las bases bioquímicas de la ingeniería genética y la biotecnología, se revisaran generalidades de cada una, pero principalmente su aplicación a problemas relacionados con el avance científico y el desarrollo de la sociedad. Se analizaran aspectos bioéticas.
CONTENIDO:
1. Bases de la bioquímica y Biología molecular.
* Metabolismo
* Catabolismo
* Anabolismo
* Metabolismo de las principales moléculas orgánicas.
* Dogma central de la Biología molecular
2. Importancia de la biotecnología.
* Enzimas de restricción
* Clonación de genes.
* Técnicas de recombinación.
* Hibridación de ácidos nucleídos.
* Terapia génica.
3. Aspectos bioéticos de la biotecnología
* Clonación
* Fecundación Invitro
* Trasplante de órganos
4. Importancia de la biotecnología.
* Microbiología industrial
* Tecnología de la fermentación
* Ingeniería enzimática
* Producción de antibióticos
INTRODUCCIÓN.
Bioquímica
La bioquímica estudia la base molecular de la vida. En los procesos vitales interaccionan un gran número de substancias de alto peso molecular o macromoléculas con compuestos de menor tamaño, dando por resultado un número muy grande de reacciones coordinadas que producen la energía que necesita la célula para vivir, la síntesis de todos los componentes de los organismos vivos y la reproducción celular.
Al conjunto de reacciones que suceden dentro de los seres vivos se le llama metabolismo.
Actualmente se conoce a detalle la estructura tridimensional de las macromoléculas de mayor importancia biológica, los ácidos nucleicos y las proteínas, lo que ha permitido entender a nivel molecular sus funciones biológicas.
Gracias al conocimiento de la estructura de los ácidos nucleicos, se esclarecieron los mecanismos de transmisión de la información genética de generación a generación, y también los mecanismos de expresión de esa información, la cual determina las propiedades y funciones de las células, los tejidos, los órganos y los organismos completos.
Conocer a detalle la estructura de varias proteínas ha sido muy útil en la elucidación de los mecanismos de las reacciones enzimáticas. Prácticamente todas las reacciones que integran el metabolismo son reacciones enzimáticas.
El tipo de especie química y los mecanismos de acción que intervienen en el almacenamiento, replicación y transferencia de la información genética, así como las reacciones que forman el metabolismo son prácticamente idénticas, desde las bacterias hasta los organismos superiores. No todas las células contienen y expresan la misma información, pero las reacciones que sí llevan a cabo, utilizan enzimas practicamente idénticas. De hecho las diferencias y similitudes entre ellas se han utilizado para establecer la secuencia de aparición de las especies. Los virus tienen algunas variantes, por ejemplo; los cromosomas de los retrovirus están constituidos por moléculas de ARN y en algunos fagos (virus que atacan a las bacterias) tienen ADN de una sola cadena. Los virus no cuentan con un metabolismo que les permita vivir en forma autónoma, sólo se pueden reproducir y expresarse dentro de las células que invaden.
Las reacciones que constituyen el metabolismo están localizadas en determinadas estructuras celulares que forman unidades discretas que se llaman organelos. Las reacciones se llevan a cabo en los lugares en donde se encuentran las enzimas que las catalizan. La célula no es un saco sin estructura, sino que es un sistema muy complejo y altamente organizado.
Ingeniería Genética y Biotecnología
A nivel básico la biotecnología se puede definir como una técnica que utiliza células vivas, cultivo de tejidos o moléculas derivadas de un organismo como las enzimas para obtener o modificar un producto, mejorar una planta o animal o desarrollar un microorganismo para utilizarlo con un propósito específico.
La diferencia aportada por la biotecnología moderna es que actualmente el hombre no sólo sabe cómo usar las células u organismos que le ofrece la naturaleza, sino que ha aprendido a modificarlos y manipularlos en función de sus necesidades. La biotecnología tal como la conocemos actualmente empezó en los años 50 con el descubrimiento por James Watson y Francis Crick de la estructura de la molécula de ADN* (ácido desoxirribonucleico) que es donde se almacena la información genética (la herencia) en todos los seres vivos.
En contra de lo que pueda parecer, la Biotecnología no es un campo nuevo de actividad empresarial, su desarrollo puede remontarse a varios miles de años atrás cuando el hombre aprendió a producir pan y otros productos como el queso, la cerveza y el vino.
El hombre lleva varios miles de años modificando los vegetales que utiliza como alimento. Por ejemplo, las repollitos de Bruselas, la coliflor y el brócoli son variedades artificiales de la misma planta (aunque no lo parezcan). Lo mismo se puede decir de las decenas de variedades de manzanas, maíz, papas, trigo, entre otros. Los antecedentes salvajes de muchas de estas plantas, cuando existen, son tan poco parecidas que no serían reconocidos como tales por alguien que no fuera experto.
En cuanto a la "mezcla de especies", el triticale, un híbrido de trigo y centeno, lleva décadas prosperando en terrenos de mala calidad (útiles para centeno, pero no para trigo), pero con algunas buenas propiedades del trigo, lo que lo hace mucho más valioso para alimentación humana.
Sin embargo, la ingeniería genética permite ahora llevar a cabo, en pocos años y de forma controlada, lo que antes podía costar décadas o siglos, o conseguir efectos que sólo estaban en los sueños de los agricultores, pero que eran imposibles con las viejas técnicas de cruce y selección.La ingeniería genética se utilizó inicialmente (por su alto coste) para producir sustancias de usos farmacéutico, como la insulina, modificando genéticamente microorganismos. Con los posteriores desarrollos, se obtuvieron también enzimas para uso industrial, como la quimosina recombinante, utilizada, al igual que la obtenida de estómagos de terneros jóvenes (su fuente original, el "cuajo"), para elaborar el queso. Posteriormente se han obtenido vegetales (y animales) modificados genéticamente para mejorar sus propiedades.Los productos de la biotecnología están alrededor nuestro. El yogurt, la cerveza, el vino y el queso de nuestra heladera son productos de la biotecnología. Los pickles, el pan, y el vinagre de nuestra cocina también lo son.Cientos de años atrás, la gente fue descubriendo, casi por accidente, cómo hacer uso de los procesos biológicos que ocurren dentro de las células vivientes. Sin entender los procesos, podían ver los resultados. Descubrieron, por ejemplo, que ciertos microorganismos, como las bacterias y los hongos podían producir vinagre, cerveza o vino cuando crecían en grandes tinas. Estos procesos fueron llamados fermentación. A través de prueba y error, aprendieron el control de estos procesos y a producir grandes cantidades de un amplio rango de productos.
BIBLIOGRAFIA:
* Curtis, H., Biología. México, Médica-Panamericana, i 983.
* Díaz Zagoya, J. C., Bioquímica. México, Interamericana-McGraw-Hill, 1995.
* Gardner, E. J., Principios de Genética. México, Limusa,. Wiley, 1975.
CONTENIDO
- Metabolismo.
El metabolismo es el conjunto de reacciones bioquímicas y procesos físico-químicos que ocurren en una célula y en el organismo. Éstos complejos procesos interrelacionados son la base de la vida a escala molecular, y permiten las diversas actividades de las células: crecer, reproducirse, mantener sus estructuras, responder a estímulos, etc.
El metabolismo se divide en dos procesos conjugados: catabolismo y anabolismo. Las reacciones catabólicas liberan energía; un ejemplo es la glucólisis, un proceso de degradación de compuestos como la glucosa, cuya reacción resulta en la liberación de la energía retenida en sus enlaces químicos. Las reacciones anabólicas, en cambio, utilizan esta energía liberada para recomponer enlaces químicos y construir componentes de las células como lo son las proteínas y los ácidos nucleicos. El catabolismo y el anabolismo son procesos acoplados que hacen al metabolismo en conjunto, puesto que cada uno depende del otro.
La economía que la actividad celular impone sobre sus recursos obliga a organizar estrictamente las reacciones químicas del metabolismo en vías o rutas metabólicas, donde un compuesto químico (sustrato) es transformado en otro (producto), y este a su vez funciona como sustrato para generar otro producto, siguiendo una secuencia de reacciones bajo la intervención de diferentes enzimas (generalmente una para cada sustrato-reacción). Las enzimas son cruciales en el metabolismo porque agilizan las reacciones físico-químicas, pues hacen que posibles reacciones termodinámicas deseadas pero "desfavorables", mediante un acoplamiento, resulten en reacciones favorables. Las enzimas también se comportan como factores reguladores de las vías metabólicas, modificando su funcionalidad –y por ende, la actividad completa de la vía metabólica– en respuesta al ambiente y necesidades de la célula, o según señales de otras células.
El metabolismo de un organismo determina qué sustancias encontrará nutritivas y cuáles encontrará tóxicas. Por ejemplo, algunas procariotas utilizan sulfuro de hidrógeno como nutriente, pero este gas es venenoso para los animales.La velocidad del metabolismo, el rango metabólico, también influye en cuánto alimento va a requerir un organismo.
Una característica del metabolismo es la similitud de las rutas metabólicas básicas incluso entre especies muy diferentes. Por ejemplo: la secuencia de pasos químicos en una vía metabólica como el ciclo de Krebs es universal entre células vivientes tan diversas como la bacteria unicelular Escherichia coli y organismos pluricelulares como el elefante.3 Esta estructura metabólica compartida es probablemente el resultado de la alta eficiencia de estas rutas, y de su temprana aparición en la historia evolutiva.


- Anabolismo.
El anabolismo es el conjunto de procesos metabólicos constructivos en donde la energía liberada por el catabolismo es utilizada para sintetizar moléculas complejas. En general, las moléculas complejas que dan lugar a estructuras celulares son construidas a partir de precursores simples. El anabolismo involucra tres facetas. Primero, la producción de precursores como aminoácidos, monosacáridos, isoprenoides y nucleótidos; segundo, su activación en reactivos usando energía del ATP; y tercero, el conjunto de estos precursores en moléculas más complejas como proteínas, polisacáridos, lípidos y ácidos nucleicos.
Los organismos difieren en cuántas moléculas pueden sintetizar por sí mismos en sus células. Los organismos autótrofos, como las plantas, pueden construir moléculas orgánicas complejas y proteínas por sí mismos a partir moléculas simples como dióxido de carbono y agua. Los organismos heterótrofos, en cambio, requieren de una fuente de sustancias más complejas, como monosacáridos y aminoácidos, para producir estas moléculas complejas. Los organismos pueden ser clasificados por su fuente de energía:
Fotoautótrofos y fotoheterótrofos, que obtienen la energía del Sol.
Quimioheterótrofos y quimioautótrofos, que obtienen la energía mediante reacciones oxidativas.
Fijación del carbono
Células de plantas (rodeadas por paredes violetas) y dentro, cloroplastos, donde se da la fotosíntesis.
La fotosíntesis es la síntesis de glucosa a partir de energía solar, dióxido de carbono (CO2) y agua (H2O), con oxígeno como producto de desecho. Este proceso utiliza el ATP y el NADPH producido por los centros de reacción fotosintéticos para convertir el CO2 en 3-fosfoglicerato, que puede ser convertido en glucosa. Esta reacción de fijación del CO2 es llevada a cabo por la enzima RuBisCO como parte del ciclo de Calvin.62 Se dan tres tipos de fotosíntesis en las plantas; fijación del carbono C3, fijación del carbono C4 y fotosíntesis CAM. Estos difieren en la vía que el CO2 sigue en el ciclo de Calvin, con plantas C3 que fijan el CO2 directamente, mientras que las fotosíntesis C4 y CAM incorporan el CO2 en otros compuestos primero como adaptaciones para soportar la luz solar intensa y las condiciones secas.
En procariotas fotosintéticas, los mecanismos de la fijación son más diversos. El CO2 puede ser fijado por el ciclo de Calvin, y asimismo por el Ciclo de Krebs inverso,o la carboxilación del acetil-CoA. Los quimioautótrofos también pueden fijar el CO2 mediante el ciclo de Calvin, pero utilizan la energía de compuestos inorgánicos para llevar a cabo la reacción.

- Catabolismo.
El catabolismo es el conjunto de procesos metabólicos que liberan energía. Estos incluyen degradación y oxidación de moléculas de alimento, así como reacciones que retienen la energía del Sol. El propósito de estas reacciones catabólicas es proveer energía, poder reductor y componentes necesitados por reacciones anabólicas. La naturaleza de estas reacciones catabólicas difiere de organismo en organismo. Sin embargo, estas diferentes formas de catabolismo dependen de reacciones de reducción-oxidación que involucran transferencia de electrones de moléculas donantesbólicas es proveer energía, poder reductor y componentes necesitados por reacciones anabólicas. La naturaleza de estas reacciones catabólicas difiere de organismo en organismo. Sin embargo, estas diferentes formas de catabolismo dependen de reacciones de reducción-oxidación que involucran transferencia de electrones de moléculas donantes (como las moléculas orgánicas, agua, amoníaco, sulfuro de hidrógeno e iones ferrosos), a aceptores de dichos electrones como el oxígeno, el nitrato o el sulfato.
En los animales, estas reacciones conllevan la degradación de moléculas orgánicas complejas a otras más simples, como dióxido de carbono y agua. En organismos fotosintéticos como plantas y cianobacteria, estas transferencias de electrones no liberan energía, pero son usadas como un medio para almacenar energía solar. El conjunto de reacciones catabólicas más común en animales puede ser separado en tres etapas distintas. En la primera, moléculas orgánicas grandes como las proteínas, polisacáridos o lípidos son digeridos en componentes más pequeños fuera de las células. Luego, estas moléculas pequeñas son llevadas a las células y convertidas en moléculas aún más pequeñas, generalmente acetilos que se unen covalentemente a la coenzima A, para formar la acetil-coenzima A, que libera energía. Finalmente, el grupo acetil en la molécula de acetil CoA es oxidado a agua y dióxido de carbono, liberando energía que se retiene al reducir la coenzima nicotinamida adenina dinucleótido (NAD+) en NADH.
El catabolismo de carbohidratos es la degradación de los hidratos de carbono en unidades menores. Los carbohidratos son usualmente tomados por la célula una vez que fueron digeridos en monosacáridos. Una vez dentro de la célula, la ruta de degradación es la glucólisis, donde los azúcares como la glucosa y la fructosa son transformados en piruvato y algunas moléculas de ATP son generadas. El piruvato o ácido pirúvico es un intermediario en varias rutas metabólicas, pero la mayoría es convertido en acetil CoA y cedido al ciclo de Krebs. Aunque más ATP es generado en el ciclo, el producto más importante es el NADH, sintetizado a partir del NAD+ por la oxidación del acetil-CoA. La oxidación libera dióxido de carbono como producto de desecho. Una ruta alternativa para la degradación de la glucosa es la ruta pentosa-fosfato, que reduce la coenzima NADPH y produce azúcares de 5 carbonos como la ribosa, el azúcar que forma parte de los ácidos nucleicos.
Las grasas son catalizadas por la hidrólisis a ácidos grasos y glicerol. El glicerol entra en la glucólisis y los ácidos grasos son degradados por beta oxidación para liberar acetil CoA, que es luego cedido al nombrado ciclo de Krebs. Debido a sus proporciones altas del grupo metileno, los ácidos grasos liberan más energía en su oxidación que los carbohidratos, ya que los carbohidratos como la glucosa tienen más oxígeno en sus estructuras.
Los aminoácidos son usados principalmente para sintentizar proteínas y otras biomoléculas; sólo los excedentes son oxidados a urea y dióxido de carbono como fuente de energía. Esta ruta oxidativa empieza con la eliminación del grupo amino por una aminotransferasa. El grupo amino es cedido al ciclo de la urea, dejando un esqueleto carbónico en forma de cetoácido. Los aminoácidos glucogénicos pueden ser transformados en glucosa mediante gluconeogénesis.

Biomoléculas principales
- Aminoácidos y proteínas
Las proteínas están compuestas por los aminoácidos, dispuestos en una cadena lineal y unidos por enlaces peptídicos. Las enzimas son proteínas que catalizan las reacciones químicas en el metabolismo. Otras proteínas tienen funciones estructurales o mecánicas, como las proteínas del citoesqueleto que forman un sistema de andamiaje para mantener la forma de la célula. Las proteínas también son partícipes de la comunicación celular, la respuesta inmune, la adhesión celular y el ciclo celular.
- Lípidos
Los lípidos son las biomoléculas que más diversidad presentan. Su función estructural básica es formar parte de las membranas biológicas como la membrana celular, o bien como recurso energético. Los lípidos son definidos normalmente como moléculas hidrófobicas o anfipáticas, que se disuelven en solventes orgánicos como la bencina o el cloroformo. Las grasas son un grupo de compuestos que incluyen ácidos grasos y glicerol; una molécula de glicerol junto a tres ácidos grasos éster dan lugar a una molécula de triglicérido. Se pueden dar variaciones de esta estructura básica, que incluyen cadenas laterales como la esfingosina de los esfingolípidos y los grupos hidrofílicos tales como los grupos fosfato en los fosfolípidos. Esteroides como el colesterol son otra clase mayor de lípidos sintetizados en las células.
- Carbohidratos
Los carbohidratos son aldehídos o cetonas con grupos hidroxilo que pueden existir como cadenas o anillos. Los carbohidratos son las moléculas biológicas más abundantes, y presentan varios papeles en la célula; algunos actúan como moléculas de almacenamiento de energía (almidón y glucógeno) o como componentes estructurales (celulosa en las plantas,quitina en los animales). Los carbohidratos básicos son llamados monosacáridos e incluyengalactosa, fructosa, y el más importante la glucosa. Los monosacáridos pueden sintetizarse y formar polisacáridos.
- Nucleótidos
Los polímeros de ADN (ácido desoxirribonucléico) y ARN (ácido ribonucléico) son cadenas denucleótidos. Estas moléculas son críticas para el almacenamiento y uso de la información genética por el proceso de transcripción y biosíntesis de proteínas. Esta información se encuentra protegida por un mecanismo de reparación del ADN y duplicada por un mecanismo de replicación del ADN. Algunos virus tienen un genoma de ARN, por ejemplo el HIV, y utilizan retrotranscripción para crear ADN a partir de su genoma viral de ARN; estos virus son denominados retrovirus. El ARN de ribozimas como los ribosomas es similar a las enzimas y puede catabolizar reacciones químicas. Los nucleósidos individuales son sintentizados mediante la unión de bases nitrogenadas con ribosa. Estas bases son anillos heterocíclicos que contienen nitrógeno y, según presenten un anillo o dos, pueden ser clasificadas como pirimidinas o purinas, respectivamente. Los nucleótidos también actúan como coenzimas en reacciones metabólicas
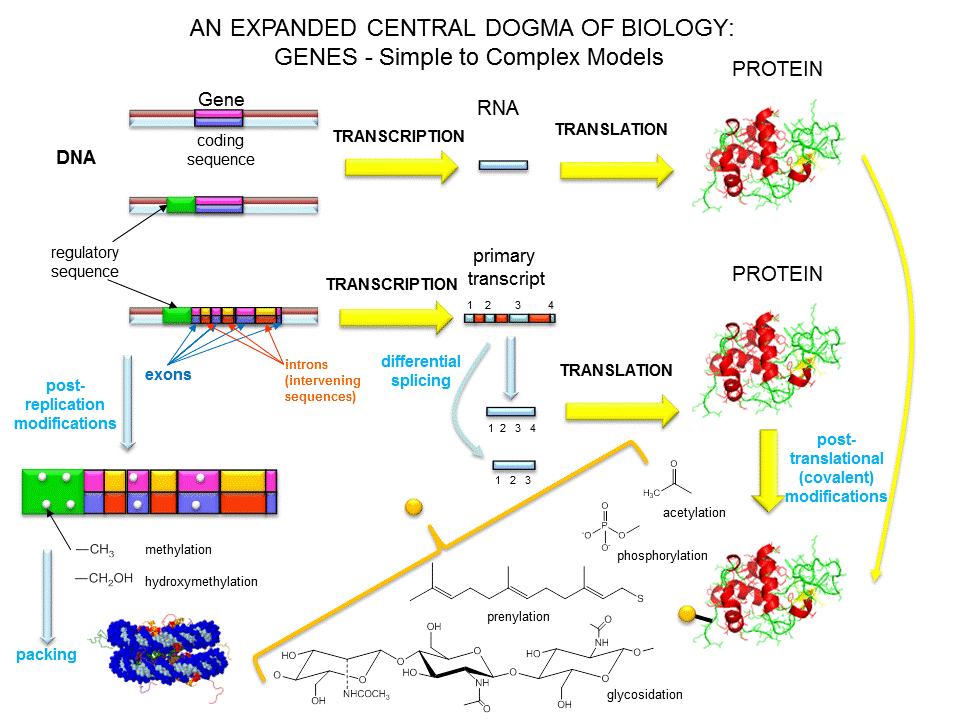
Dogma central de la Biología Molecular
Dado que en la célula cada molécula tiene una función y las proteínas son las encargadas de realizar “el trabajo duro” (formar estructuras, catalizar reacciones enzimáticas, activar genes, entre otras), la información contenida en forma de genes debe, de alguna manera, ser convertida en proteínas. En este apartado y en el próximo describimos los procesos que involucran la traducción de este código y la síntesis de proteínas.
La información genética está contenida en los genes, segmentos de ADN que llevan información para fabricar un producto funcional determinado. Nuestro genoma tiene aproximadamente 30.000 genes. Sólo una pequeña parte del genoma es codificante; la mayor parte corresponde a secuencias cortas móviles no codificantes o a secuencias regulatorias.
Para que la información pase de una molécula a otra, primero debe copiarse, en un proceso que se llama replicación y que ocurre en el núcleo. Pero como el ADN se encuentra en el núcleo y las proteínas son sintetizadas en el citoplasma, debe existir una molécula que funcione como intermediaria. Este papel lo cumple el ácido ribonucleico mensajero (ARNm). El ADN se copia en ARNm en el núcleo, en un proceso denominado transcripción. Luego la información contenida en el ARNm es empleada para construir proteínas en el proceso de traducción, que tiene lugar en el citoplasma.
Estos tres procesos secuenciales constituyen el llamado dogma central de la Biología, que establece que la información fluye desde el ADN al ARN y de este a las proteínas. (Además, las proteínas controlan el proceso de replicación del ADN uniéndose a una secuencia específica en el ADN. De esta manera pueden activar o inhibir la transcripción de un gen determinado.)
BIBLIOGRAFIA:
* Curtis, H., Biología. México, Médica-Panamericana, i 983.
* Díaz Zagoya, J. C., Bioquímica. México, Interamericana-McGraw-Hill, 1995.
* Gardner, E. J., Principios de Genética. México, Limusa,. Wiley, 1975.
EXPOSICIONES.
EXPOSICIÓN 1. Transformación Plasmidos y Endonucleacidos de Restricción.
Los plásmidos, vectores o también llamados plasmidios, son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb . El número de plásmidos puede variar, dependiendo de su tipo, desde una sola copia hasta algunos cientos por célula. El término plásmido fue presentado por primera vez por el biólogo molecular norteamericano Joshua Lederberg en 1952.
Las moléculas de ADN plásmidico, adoptan una conformación tipo doble hélice al igual que el ADN de los cromosomas, aunque, por definición, se encuentran fuera de los mismos. Se han encontrado plásmidos en casi todas las bacterias. A diferencia del ADN cromosomal, los plásmidos no tienen proteínas asociadas.
En la mayoría de los casos se considera genético dispensable. Sin embargo, posee información genética importante para las bacterias. Por ejemplo, los genes que codifican para las proteínas que las hace resistentes a los antibióticos están, frecuentemente, en los plásmidos.
Hay algunos plásmidos integrativos, es decir, que tienen la capacidad de insertarse en el cromosoma bacteriano. Estos rompen momentáneamente el cromosoma y se sitúan en su interior, con lo cual, automáticamente la maquinaria celular también reproduce el plásmido. Cuando ese plásmido se ha insertado se les da el nombre de episoma.
Los plásmidos se utilizan en ingeniería genética por su capacidad de reproducirse de manera independiente del ADN cromosomal como así también por que es relativamente fácil manipularlos e insertar nuevas secuencias genéticas.
Los plásmidos usados en Ingeniería Genética suelen contener uno o dos genes que les confieren resistencia a antibióticos y permiten seleccionar clones recombinantes. Hay otros métodos de selección además de la resistencia a antibióticos, como los basados en fluorescencia o en proteínas que destruyen las células sin uso de antibióticos. Estos nuevos métodos de selección de plásmidos son de uso frecuente en agrobiotecnología, debido a la fuerte crítica de grupos ecologistas contra la posibilidad de presencia de antibióticos en los organismos modificados genéticamente.

EXPOSICIÓN 2. Enzimas de Restriccion
Las enzimas de restricción, también conocidas como endonucleasas, son enzimas que cortan los enlaces fosfodiester del material genético a partir de una secuencia que reconocen.Las mismas permiten cortar DNA de hebra doble, donde reconocen secuencias palindrómicas (secuencias que se leen igual en ambas direcciones)
Son extraídas de organismos procarióticos (bacterias), donde actúan como un mecanismo de defensa, para degradar material genético extraño que entre en la célula. Las bacterias tienen la capacidad de metilar su DNA, lo cual sirve para distinguir entre el DNA extraño y el DNA propio. Las enzimas de restricción no pueden cortar DNA metilado, de este modo solo afectan el DNA extranjero y no el DNA bacterial.
Existen 3 tipos de enzimas de restricción:
1. Tipo I y Tipo III:
a. Tienen actividad de restricción (cortan) y modificación (metilan).
b. Cortan a cierta distancia de la secuencia de reconocimiento, las Tipo I cortan lejos de la secuencia de reconocimiento, ya sea río arriba o río abajo. Las Tipo III cortan de 5-8 bases antes o despúes de la secuencia que reconocen.
c. Necesitan ATP para moverse a través de la molécula de DNA, desde el lugar de reconocimiento hasta el sitio del corte.
2. Tipo II:
a. Sólo tienen actividad de restricción.
b. Cortan de manera consistente y predecible dentro de la secuencia que reconocen.
c. Sólo requieren Mg++ como cofactor.
d. No necesitan ATP.
Aplicaciones de las enzimas de restricción:
1. Hacer mapa de restricción de un plásmido o bacteriófago.
2. Fragmentar DNA genómico para separación de electroforesis y “Southern Blot”.
3. Generación de fragmentos para ser usados como sondas marcadas en “Southern”y “Northern” blotting.
4. Generación de fragmentos para ser subclonados en los vectores apropiados, creación de DNA recombinante
Las endonucleasas se nombran a partir de las bacterias de las que son extraídas, su nombre está dado según el género y la especie de la bacteria de donde se aisló por primera vez esta enzima. La primera letra representa el género de la bacteria, las próximas dos indican la especie, una cuarta letra indica la cepa, y un número al final indica la cantidad de enzimas que se han aislado de esa cepa. Ej:
Eco RI à E = género Escherichia
co = especie coli
R = cepa RV 13
I = primera endonucleasa aislada de esta cepa
Una enzima de restricción (o endonucleasas de restricción) es aquella que puede reconocer una secuencia característica de nucleótidos dentro de una molécula de ADN y cortar el ADN en ese punto en concreto, llamado sitio o diana de restricción, o en un sitio no muy lejano a éste, dependiendo de la enzima. Los sitios de restricción cuentan con entre 4 y 12 pares de bases, con las que son reconocidos.
El mecanismo de corte de DNA se realiza a través de la ruptura de dos enlaces fosfodiéster en la doble hebra, lo que da lugar a dos extremos de DNA. Éstos pueden ser romos (cuando los enlaces rotos coinciden) o Cohesivos/escalonados. Estos últimos tienen tendencia a volver a unirse de modo espontáneo, ya que los extremos se pueden unir a otros extremos coincidentes que pueda haber en la cercanía (Apareamiento de Watson & Crick).
Los fragmentos de ADN obtenidos de este modo pueden unirse por otras enzimas llamadas ligasas. Conocemos así el ADN vector, que sería aquel que es capaz de replicarse independientemente del ADN de la célula anfitriona en la cual crece. Dentro de este grupo de vectores están los plásmidos, moléculas circulares de ADN halladas en las bacterias.
Las enzimas de restricción que a pesar de ser distintas y provenir de distintas especies, tienen la misma secuencia de reconocimiento y dejan el mismo extremo cohesivo, pero no cortan en el mismo sitio, son llamadas isoesquizómeros. Por ejemplo, están los isoesquizómeros Asp718 y KpnI.
Uno de los campos en los que el uso de enzimas de restricción ha tenido mayor implicación ha sido el diagnóstico de enfermedades genéticas relacionadas con cambios en la secuencia del ADN, ya sean mutaciones puntuales, inserciones o deleciones de fragmentos. Si éstas se producen en un sitio de reconocimiento de la enzima de restricción, al producirse eliminarán o agregarán nuevos sitios de corte. Al aplicar esta enzima al gen de una persona sana y una enferma se deberían observar distintas cantidades de fragmentos para cada caso en una electroforesis.


EXPOSICION 3. Ingeniería Genética.
La ingeniería genética es una parte de la biotecnología que se basa en la manipulación genética de organismos con un propósito predeterminado, aprovechable por el hombre: se trata de aislar el gen que produce la sustancia e introducirlo en otro ser vivo que sea más sencillo de manipular. Lo que se consigue es modificar las características hereditarias de un organismo de una forma dirigida por el hombre, alterando su material genético. El proceso puede utilizarse ya en bacterias y en células eucariotas vegetales o animales. Una vez adicionada o modificada la carga cromosómica, el organismo en cuestión sintetiza la proteína deseada y el aumento del rendimiento de la producción puede obtenerse mediante el aumento en la población portadora. Las bases de la ingeniería genética han consistido en resolver el problema de la localización e inserción de genes y la multiplicación redituable de las factorías logradas. Las técnicas utilizadas por la ingeniería genética son varias, y cada una atiende un aspecto de la tarea de preparación y solución de los problemas específicos de esta tecnología, sin embargo muchas de ellas ha tenido éxito en otros campos tecno científicos.
APLICACIONES DE LA INGENIERÍA GENÉTICA
La aplicación de las técnicas utilizadas por la Ingeniería Genética ha permitido elevar la calidad de vida del ser humano. Los organismos transgénicoshan pasado a ocupar una posición central en la biotecnología moderna, porque permiten hacer modificaciones muy específicas del genoma que vale la pena analizar con detalle, debido a sus importantes aplicaciones presentes y futuras.
Obtención de proteínas de interés médico y económico
- antibióticos
- enzimas
- hormonas: insulina, hormona del crecimiento, eritropoyetina
- vacunas
- proteínas sanguíneas: seroalbúmina, factores de coagulación…
Con el mejoramiento genético de los vegetales, se espera conseguir:
- Mayor adaptación a diversos ambientes.
- Mejores características agronómicas (resistencia, desgrane, buena cobertura, etc.).
- Resistencia a plagas y enfermedades.
- Resistencia a la sequía, temperaturas bajas o altas, etc.
Para incrementar la calidad de los productos se persigue:
- Alto valor nutritivo (proteínas y vitaminas).
- Mayor coloración, sabor y/o tamaño de los frutos.
- Resistencia al transporte y almacenamiento.
- Reducción de la cantidad de ciertas sustancias indeseables en los productos, etc.
Obtención de animales y vegetales transgénicos
Animales
- obtención de órganos animales (cerdos) con genes humanos para no ser rechazados en trasplantes.
- animales con carnes y huevos con menos colesterol y grasas
- pollos sin plumas
Vegetales
- resistentes a insectos: maíz y algodón con un gen que produce una toxina para orugas y escarabajos
- resistentes a herbicidas: soja, algodón, maíz, resisten a altas concentraciones de herbicidas que se echan en los campos para erradicar malas hierbas
- resistentes a condiciones ambientales: frío, sequía, altasalinidad, etc.
Inconvenientes
Los expertos advierten que detrás de estas mejoras y nuevas aplicaciones se esconden también riesgos y peligros de notable importancia.
Como sucede siempre, las desventajas provienen o pueden proceder del mal uso de las técnicas mencionadas, lo cual es motivo de preocupación por los riesgos e implicaciones que pueden derivarse. A ello ha dado respuesta el Comité Internacional de Bioética de la Unesco fijando unos objetivos que pueden concretarse en dos:
a) evitar aspectos del progreso que atenten contra la dignidad humana
b) que las posibilidades científicas no generen peligrosidad por falta de definiciones éticas.
Los criterios para evitar dichos inconvenientes establecen una serie de limitaciones por motivos ecológicos, sanitarios, morales, sociales, políticos... y en concreto se trata sobre todo de la salvaguarda de la dignidad y los derechos humanos, de no dar posibilidad a la discriminación social ni ideológica de evitar desastres ecológicos y de impedir el desarrollo o aparición de enfermedades que pudieran ser incontrolables.
- Uno de estos peligros es el hecho de que detrás de los proyectos de manipulación genética están las compañías multinacionales muy preocupadas por el interés económico.
- También pueden “contaminar” otras plantas no transgénicas.
- Pueden llegar a ser cancerígenas en el caso de ser consumidos por sujetos proclives o en un estado inmunológico deficiente. No obstante esto es una hipótesis pero que muchos médicos que están en contra de los alimentos transgénicos lo afirman.
- Puede producir alergias, algo que preocupa mucho a los productores de estos alimentos. Puede ser debida al material genético transferido, a la formación inesperada de un alérgeno o a la falta de información sobre la proteína que codifica el gen insertado.

EXPOSICIÓN 4. VECTORES.






EXPOSICIÓN 5. Clonacion de Genes.
Clonación de genes, el proceso mediante el cual puede aislarse un gen de entre todos los genes diferentes que existen en un organismo, lo que permite realizar su caracterización. Esto se consigue con la preparación de una batería de bacterias que contienen todos los genes distintos presentes en un organismo de manera que cada una de ellas contiene un solo gen. Esto se lleva a cabo efectuando cortes del ADN de un individuo. Otra alternativa es la de crear un conjunto de todas las secuencias de ADN expresadas en una célula específica mediante la producción de copias complementarias de ADN a partir del ARNm hallado en dichas células (v ase Biología molecular). En ambos casos, los fragmentos de ADN se unen a un vector, un virus bacteriano conocido como bacteriófago o a un ADN circular denominado plasmido, que se introduce en una bacteria de forma que cada una adquiere solo una copia del vector y por tanto recibe solo un fragmento de ADN.
Los grupos preparados de esta forma se pueden examinar para identificar la bacteria que contiene el gen objeto de estudio. Entonces, se toma esta bacteria y se hace crecer para producir un clon de bacterias idénticas. Como el vector que contiene el ADN insertado se replica siempre que la célula bacteriana se divide, se produce la cantidad suficiente de ADN insertado clonado necesaria para caracterizar el gen. De esta manera es posible estudiar los genes que codifican proteínas que tienen un interés especial, o aquellos cuya inactivación, consecuencia de una mutación, origina una enfermedad específica. Por ejemplo, podemos determinar su secuencia y la naturaleza de la mutación que da lugar a una enfermedad.
Con posterioridad, el gen se puede expresar en la célula bacteriana para producir la proteína específica que se puede emplear en el tratamiento de enfermedades como la diabetes mellitus (insulina) o el enanismo (hormona del crecimiento). Recientemente, se han podido introducir genes funcionales clonados en los individuos, para tratar una enfermedad de forma más directa. Es probable que el empleo de estos procedimientos de tratamiento genético con ADN clonado aumente en el futuro.
La clonación (derivado del griego κλων, que significa "retoño") puede definirse como el proceso por el que se consiguen copias idénticas de un organismo, célula o molécula ya desarrollado de forma asexual.
Se deben tomar en cuenta las siguientes características:
- En primer lugar se necesita clonar las moléculas ya que no se puede hacer un órgano o parte del "clon" si no se cuenta con las moléculas que forman a dicho ser, aunque claro para hacer una clonación necesitamos saber qué es lo que buscamos clonar (ver clonación molecular).
- Ser parte de un animal ya desarrollado, porque la clonación responde a un interés por obtener copias de un determinado animal que nos interesa, y sólo cuando es adulto conocemos sus características.
- Por otro lado, se trata de crearlo de forma asexual. La reproducción sexual no nos permite obtener copias idénticas, ya que este tipo de reproducción por su misma naturaleza genera diversidad.
EXPOSICIÓN 6. Secuenciacion de DNA.
El análisis más detallado de la estructura del ADN consiste en averiguar la secuencia de nucleótidos. A lo largo del tiempo se han desarrollado diferentes métodos para obtener la secuencia de nucleótidos del ADN, sin embargo, actualmente los métodos más utilizados son el de secuenciación automática y el método enzimático de terminación de cadena de Sanger también conocido por el método didesoxi.
Método enzimático de terminación de cadena o método didesoxi de Sanger.
Para obtener la secuencia de bases nitrogenadas de un segmento de ADN por el método enzimático de terminación de cadena, se necesitan los siguientes compuestos:
- El ADN molde o segmento de ADN que se desea secuenciar. Para poder secuenciar un segmento de ADN, previamente se necesita tener gran cantidad de ese fragmento, y por tanto, hay que clonarlo en un vector apropiado. Además, debe estar en estado de hélice sencilla.
- Un enzima que replique el ADN, normalmente la ADN Polimerasa I del bacteriofago T4. La ADN Polimersa I del fago T4 emplea como molde ADN de hélice sencilla y siguiendo las reglas de complementaridad de las bases nitrogenadas va añadiendo nucleótidos a partir de un cebador o "primer".
- Un cebador o "primer" que suele ser un oligonucleótido corto de alrededor de 20 bases de longitud necesario para que la ADN polimerasa I comience a añadir nucleótidos por el extremo 3' OH. Este cebador debe poseer una secuencia de bases complementaria a la del fragmento de ADN que se desea secuenciar. Debido a que la secuencia de nucleótidos del segmento que se quiere secuenciar es desconocida, se emplea un "primer" con secuencia complementaria al vector empleado para clonar el fragmento de ADN, además, este cebador procede de una región del vector muy cercana al punto de inserción del ADN problema cuya secuencia se conoce. El "primer" utilizado suele marcarse radiactivamente.
- Los cuatro nucleótidos trifosfato (dATP, dCTP, dGTP y dTTP). A veces en vez de marcar radiactivamente el cebador, se marca radiactivamente uno de los cuatro nucleótidos trifosfato en cada reacción.
- Por último, se necesitan nucleótidos didesoxi (ddATP, ddTTP, ddCTP y ddGTP). Los nucleótidos didesoxi son nucleótidos modificados que han perdido el grupo hidroxilo de la posición 3' de la desoxirribosa. Estos nucleótidos pueden incorporarse a la cadena de ADN naciente, pero no es posible que se una a ellos ningún otro nucleótido por el extremo 3'. Por tanto, una vez incorporado un nucleótido didesoxi se termina la síntesis de la cadena de ADN.

Breve descripción del método enzimático de terminación de cadena
- En primer lugar, deben realizarse en cuatro tubos diferentes, cuatro mezclas de reacción. Cada mezcla de reacción contiene los cuatro nucleótidos trifosfato (dATP, dCTP, de dTTP y dGTP), ADN polimerasa I, un cebador marcado radiactivamente y un nucleótido didesoxi, por ejemplo ddATP, a una concentración baja. El nucleótido didesoxi utilizado (ddATP en este ejemplo) competirá con su homólogo (dATP) por incorporarse a la cadena de ADN que se está sintetizando, produciendo la terminación de la síntesis en el momento y lugar donde se incorpora.
- Por este sistema, en cada mezcla de reacción se producen una serie de moléculas de ADN de nueva síntesis de diferente longitud que terminan todas en el mismo nucleótido y marcadas todas radiactivamente por el extremo 5' (todas contienen en el extremo 5' el cebador utilizado).

- Los fragmentos de ADN de nueva síntesis obtenidos en cada mezcla de reacción se separan por tamaños mediante electroforesis en geles verticales de acrilamida muy finos (0,5 mm de espesor) y de gran longitud (cerca de 50 cm) que permiten distinguir fragmentos de ADN que se diferencian en un solo nucleótido.Los productos de cada una de las cuatro mezclas de reacción se insertan en cuatro calles o carriles diferentes del gel.
- Una vez terminada la electroforesis, el gel se pone en contacto con una película fotográfica de autorradiografía. La aparición de una banda en una posición concreta de la autorradiografía en una de las cuatro calles nos indica que en ese punto de la secuencia del ADN de nueva síntesis (complementario al ADN molde) está la base correspondiente al nucleótido didesoxi utilizado en la mezcla de reacción correspondiente.

- Teniendo en cuenta que el ADN de nueva síntesis crece en la dirección 5' ® 3', si comenzamos a leer el gel por los fragmentos de menor tamaño (extremo 5') y avanzamos aumentando el tamaño de los fragmentos (hacia 3'), obtendremos la secuencia del ADN de nueva síntesis en la dirección 5' ® 3'.
Breve descripción del método automático de secuenciación
- La principal diferencia entre método enzimático de terminación de cadena y el método automático de secuenciación radica, en primer lugar en el tipo de marcaje. En el método automático en vez de radiactividad se utiliza fluorescencia y lo habitual es realizar cuatro mezclas de reacción, cada una con nucleótido trifosfato (dTTP) marcado con un fluorocromo distinto. Este sistema permite automatizar el proceso de manera que es posible leer al mismo tiempo los ADNs de nueva síntesis producto de las cuatro mezclas de reacción.
- La segunda diferencia radica en el sistema de detección de los fragmentos de ADN. La detección del tipo de fluorescencia correspondiente a cada reacción se lleva a cabo al mismo tiempo que la electroforesis, de manera que los fragmentos de ADN de menor tamaño que ya han sido detectados se dejan escapar del gel, permitiendo este sistema aumentar el número de nucleótidos que se pueden determinar en cada electroforesis y, por consiguiente, en cada secuenciación.

En la siguiente figura se muestra un ejemplo de una secuencia obtenida por el método automático de secuenciación. Cuando aparece la letra N significa que no ha sido posible determinar el nucleótido existente en esa posición de la secuencia.

BIBLIOGRAFIA:
* Díaz Zagoya, J. C., Bioquímica. México, Interamericana-McGraw-Hill, 1995.
* González, P. A., Biología moleculary ceMar. Barcelona, Omega, 1994
* Peña, A., Arroyo, C., Gómez Puyou, A., Tapia, R. y Villa, S., Bioquímica. México, Limusa, 1979